// article
Evidence and Perception
Stronger evidence does not raise the signal, it splits it
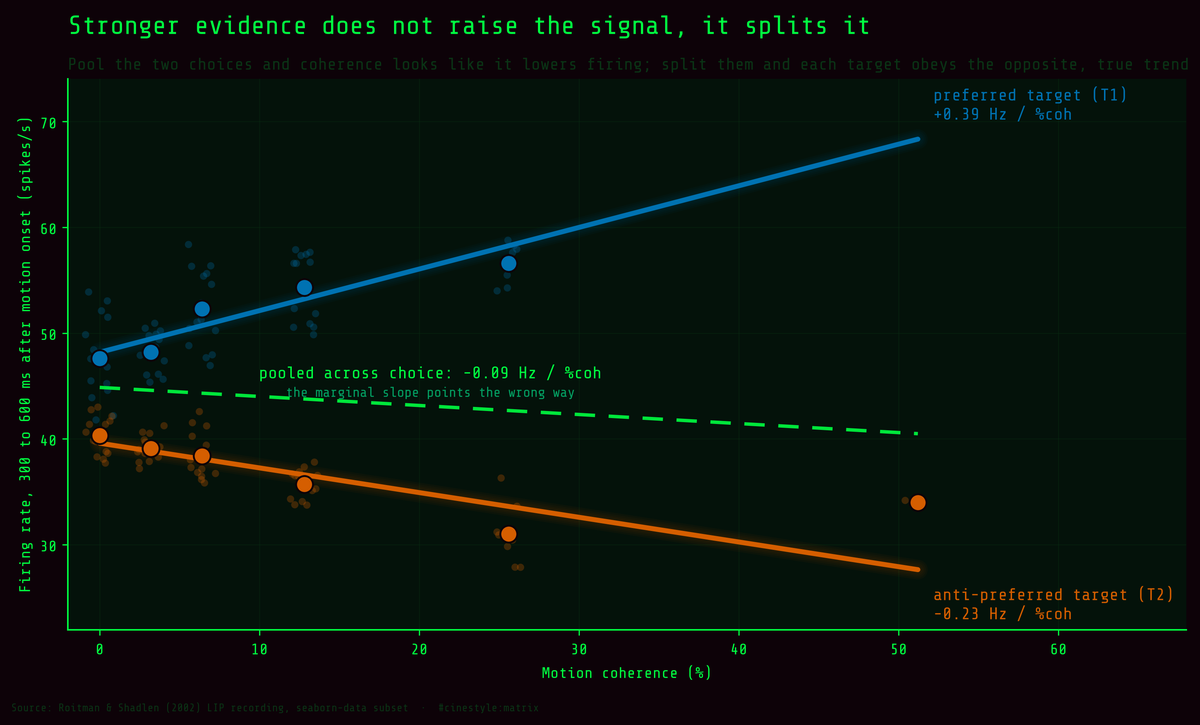
The strongest signal in a perceptual-decision recording is not the stimulus. It is the choice. This is the whole article in one frame. Pool the two choices together and coherence looks like it quietly lowers the neuron’s firing. Split the trials by which target the monkey picked and the picture reverses: each choice obeys a strong, opposite trend, and the pooled line is just the average of two things going in different directions. It is a textbook Simpson’s paradox, and you can watch it happen on a single recording.
Late in the trial, a neuron firing for the monkey’s chosen target sat at 51.1 spikes per second. A neuron firing for the rejected target sat at 37.5. That 13.6 Hz gap, measured 300 to 600 ms after the dots started moving, is the cleanest number in this whole dataset, and it has nothing to do with how strong the motion was. So I assumed when I started.
I pulled the dots set that ships with seaborn. It is a packaged demo subset of the Roitman & Shadlen 2002 random-dot motion recordings from area LIP. 848 rows, five columns: align, choice, time, coherence, firing_rate. The monkey watches a cloud of dots, some fraction moving coherently in one direction, and saccades to one of two targets to report which way. coherence is that fraction: 0, 3.2, 6.4, 12.8, 25.6, or 51.2 percent. choice is T1 or T2, the two targets. align splits the recording into two epochs: dots is locked to motion onset, sacc is locked to the eye movement. time runs from -600 to 720 ms across 67 bins. firing_rate is in spikes per second, and the units are about as arbitrary as that sounds for a demo subset of one averaged neuron.
My hypothesis going in was the obvious one. More coherence means stronger evidence, so the signal should climb with coherence. Cleaner stimulus, louder neuron.
The pooled data says the opposite. Correlate firing rate against coherence across all 848 rows and you get Pearson r = -0.13 (p = 0.0002). Negative. Average firing drops from 40.9 Hz at 0% coherence to 36.2 Hz at 51.2%. Stronger evidence, quieter neuron. That runs backwards from every textbook account of this experiment.
The pooling was lying
The trap is choice. T1 is the neuron’s preferred target, the direction its receptive field cares about. T2 is the anti-preferred one. When you average across both, you are mixing a population that should go up with coherence and one that should go down, and the mixture depends on how many trials land in each bin.
So I split it. In the late stimulus window, I fit firing rate against coherence separately for each choice:
- T1 (preferred): slope +0.39 Hz per percent coherence, r² = 0.46, p = 1.3e-10
- T2 (anti-preferred): slope -0.23 Hz per percent coherence, r² = 0.54, p = 1.5e-13
Coherence does not push one global signal up or down. It drives the two choices apart. On preferred-target trials the neuron climbs about four-tenths of a spike per second for every extra percent of coherent motion; on anti-preferred trials it sinks at a bit more than half that rate. Both effects are statistically obvious. The pooled slope is negative either way you cut it, -0.13 across all rows and -0.20 across the late-window bin means, and both are the same artifact: more high-coherence trials fall on the T2 side, so the marginal average gets dragged down even while each choice’s own slope behaves.
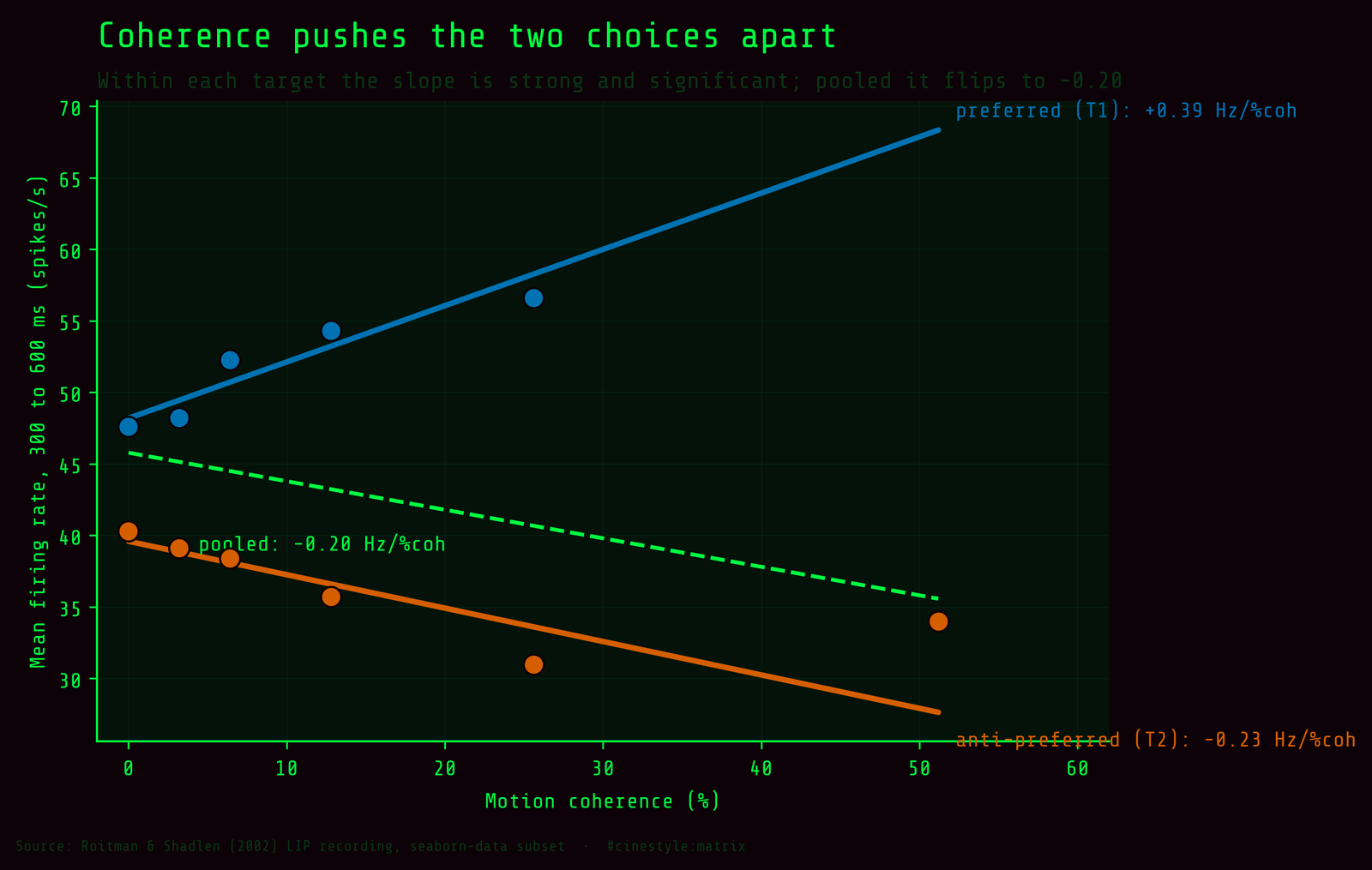
The figure shows it plainly. Two lines spreading as you move right, and the dashed grey pooled fit, slope -0.20 Hz per percent coherence through the six bin means, sloping the wrong way through the middle. One thing is off: the T1 line stops at 25.6%. In this 300 to 600 ms window there are no T1 trials at 51.2% coherence, so the highest-evidence point on the blue line is missing. The preferred target does have 51.2% trials elsewhere in the recording, 19 of them, just not in this window. That gap is part of why the pooled fit gets yanked downward at the right edge: the only 51.2% point left is the low T2 one. With a demo subset of a few hundred trials, the bins are not balanced, and you feel it here. Imbalanced bins are how a real trend hides inside a marginal that lies.
Evidence needs time to show up
Coherence is not visible the instant the dots appear. It accumulates. Picture the firing rate as a running tally that the neuron adds to with every moment of coherent motion it sees; at time zero the tally is empty no matter how strong the stimulus. I checked the early window, -80 to 80 ms around motion onset, and coherence explains essentially nothing there: r = 0.026, p = 0.79, with a total spread of 0.86 Hz across all six coherence levels. The neuron has not separated yet because there has been no time to integrate evidence.
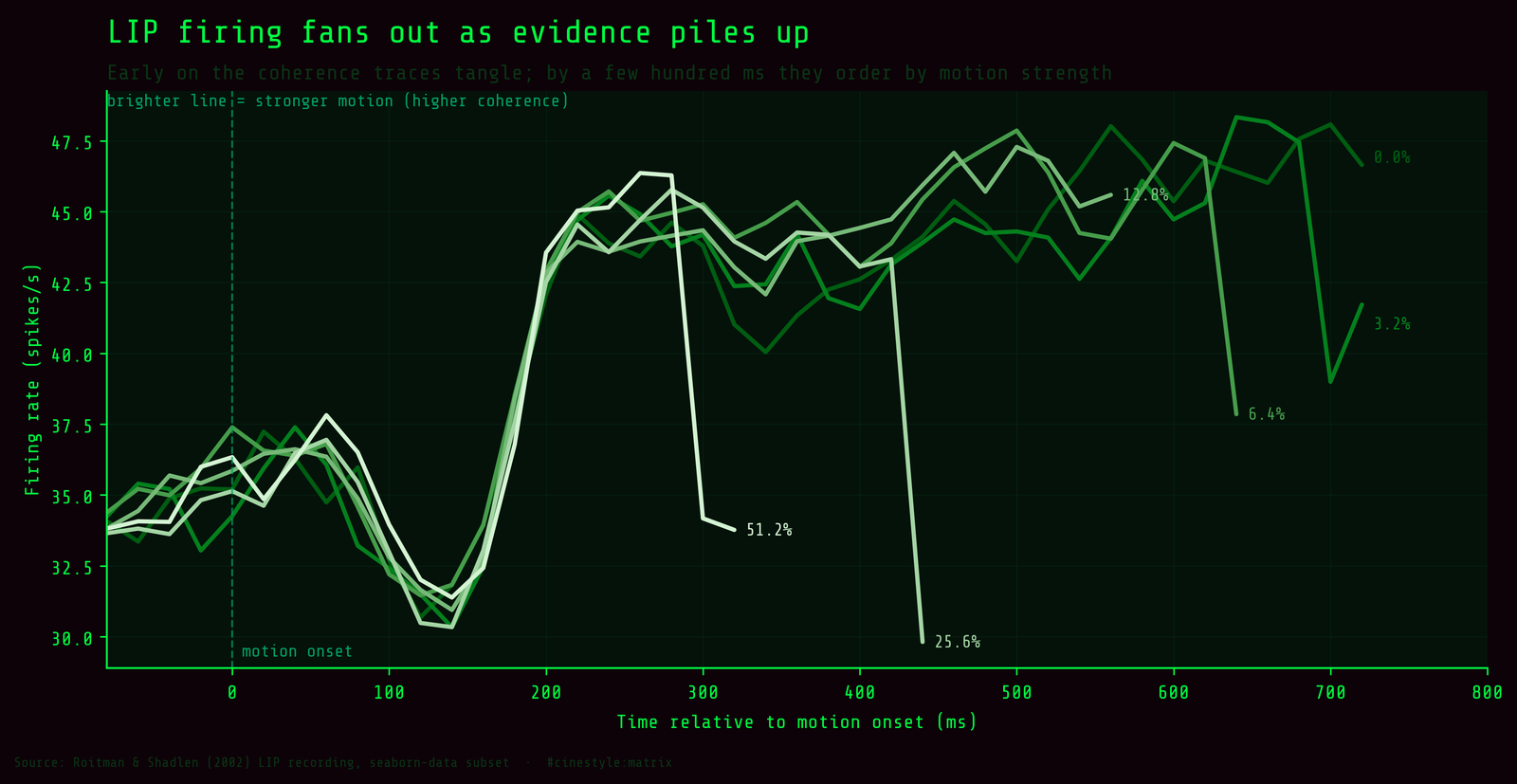
Watch the lines fan out as time runs forward. Early on they are a tangle. By a few hundred milliseconds in, the ordering is there, though it is the direction-collapsed ordering, which is why the fan in this plot looks noisier than the clean choice-split version above. This is the integration-to-bound picture people draw for LIP: the firing rate is a running tally of motion evidence, and stronger motion moves the tally faster. The signal does not arrive with the stimulus, it is built from it.
What the neuron really tracks
If coherence sets the rate of evidence, choice is the outcome. The 13.6 Hz late-window gap between T1 and T2 (Welch t = 20.7, p = 1.9e-42) dwarfs anything coherence does on its own. Lock the recording to the saccade instead of to the motion, and you can see why.
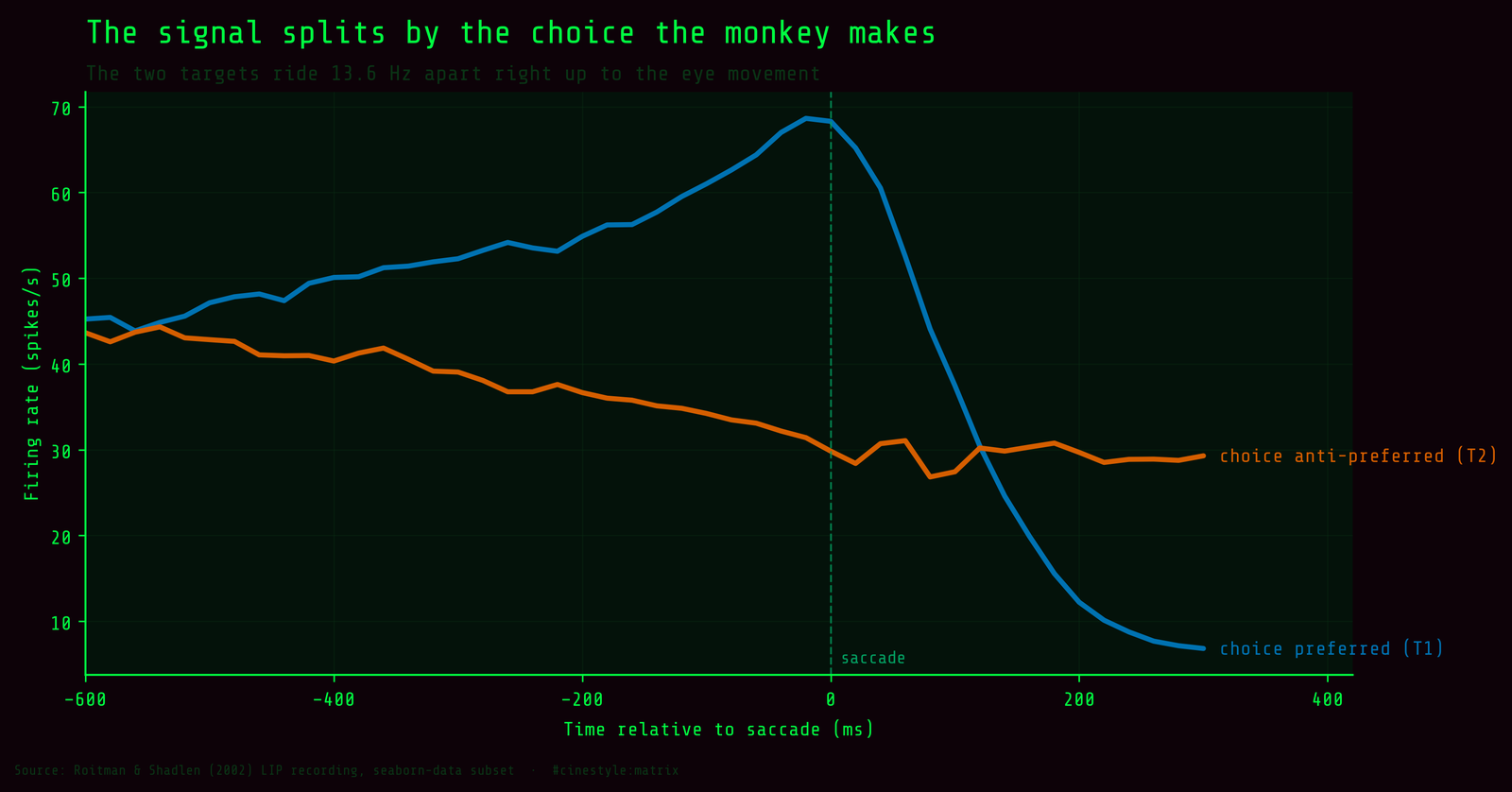
In the saccade-aligned epoch the two choice traces ride at clearly different levels right up to the eye movement at time zero. The neuron is not reporting “how strong was the motion” so much as “which target am I about to commit to.” Coherence sits upstream of that. It controls how fast and how confidently the commitment forms, but the variable the cell most cleanly encodes is the decision itself.
So the honest answer to “how cleanly does coherence drive the neural signal” is: cleanly, but only once you condition on choice, and only after evidence has had a few hundred milliseconds to pile up. Take it raw and pooled, and coherence appears to lower the firing rate, which is exactly the kind of Simpson’s-paradox mirage that makes you re-run the script before you believe your own plot. I ran it three times.
The strongest signal in a perceptual-decision recording is not the stimulus. It is the choice. The stimulus just sets how quickly the choice pulls away.